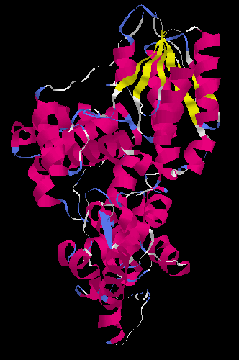|
|
1. IntroductionThe major damage caused by UV radiation of DNA is the formation of cyclobutane pyrimidine dimers (CPD), which block replication and have cytotoxic and mutagenic effects.[3] The formation of CPD has been linked to certain types of skin cancer, the most widespread form of cancer in the US. Many organisms are capable of repairing this light-induced damage by another light-dependent step, catalyzed by the enzyme DNA photolyase.[6] This enzyme binds CPD in single or double stranded DNA essentially independent of the sequence with binding constants of ~ 10-9 M. Irradiation of this enzyme-substrate complex with visible light leads to an photoinduced electron transfer from a flavine to the dimer. The resulting radical anion then splits very rapidly to form the fully functional DNA after back electron transfer to the enzyme.
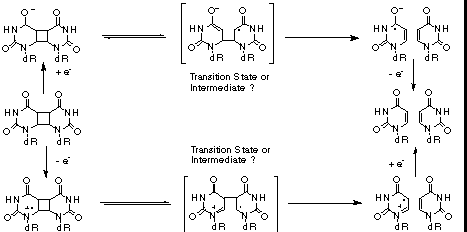
Figure 1: Ribbon drawing of the three-dimensional structure of DNA photolyase from E. coli[4] The structure of DNA photolyase from Escherichia coli has recently been determined by X-ray crystallography.[4,5] The overall structure, shown in Figure 1, consists of five parallel beta-strands, twenty alpha-helices and five short 310 helices which are organized in three major domains. One domain is formed by the beta-strands and four alpha-helices that bind the second cofactor MTHF. The other two, formed by the helical structures, are separated by a hole in the otherwise relatively flat protein surface. Based on the position of the noncovalentely bond flavine cofactor in this hole, the authors proposed that it constitutes the active site of DNA photolyase. This is also supported by the fact that the size of the opening is suitable for binding of a CPD (compare also Figure 6). The rim and one side of the binding site are positively charged (Arg226, Arg342 and Arg397), whereas the interior on the other side is relatively hydrophobic (Phe150, Val270, Trp277, Tyr281, Met345, Trp384 and Ala392). The hole lies in the middle of a trace of positive electrostatic potential on the surface, designed to bind to the phosphate backbone of DNA. The mode of binding is, however, not known since there are no data on a enzyme-substrate complex available. Although there has been much interest in the mechanism of the electron transfer catalyzed cycloreversion,[6] the details of the reaction are still unknown. Figure 2 summarizes the mechanistic possibilities. Trapping and kinetic isotope effect (KIE) studies showed that the radical cationic pathway, induced by one-electron oxidation, proceeds via a singly linked intermediate. The structure of this intermediate and the reaction energetics are, however, not known. The mechanism of the biologically more relevant reductive, radical anionic pathway is even less clear since no intermediate could be trapped and the results from KIE studies are ambiguous. [7]
Figure 2: Overall reaction scheme for the electron transfer catalyzed [2+2] cycloreversion of CPD As part of our ongoing studies of the mechanism of these important reactions, we present here an analysis of the electronic structure of the most simple CPD, the cyclobutane uracil dimer 1, the radical cation 1*+ and the radical anion 1*-. These studies represent a significant improvement over previous calculations of these species[8], since the semiempirical methods used in earlier studies are not likely to yield accurate data for radical ions. 2. Computational MethodologyThe geometries of all species discussed here were fully optimized using the Becke3LYP/6-31G* method and characterized by harmonic frequency analysis. It was shown earlier by our group[9] and others[10] that the Becke3LYP/6-31G* method gives excellent results for both neutral molecules and radical ions. The potential surfaces were calculated using single point calculations at the HF/6-31G* level of theory. Geometry optimizations and frequency analysis were performed using the GAUSSIAN94 series of programs[11] running on an IBM SP2 parallel computer at the University of Notre Dame. The potential surfaces were generated using SPARTAN 4.0[12], running on a SGI INDY R5000. Enzyme structures were visualized using RASMOL2.5.[13] |